Things might look a little different around here... Welcome to Conservation International’s new digital presence.
Things might look a little different around here... Welcome to Conservation International’s new digital presence.
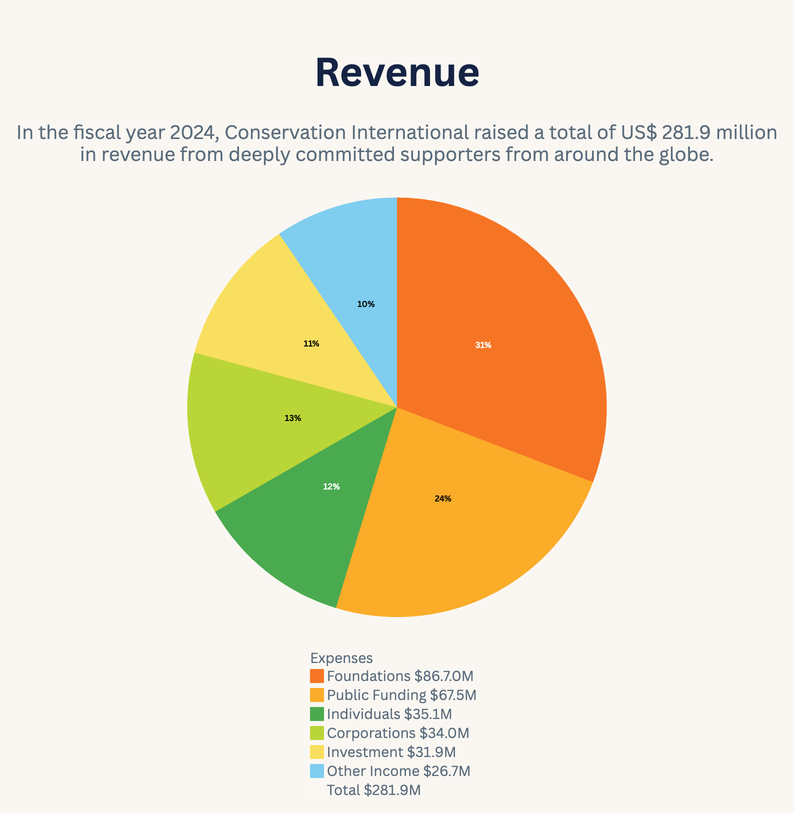
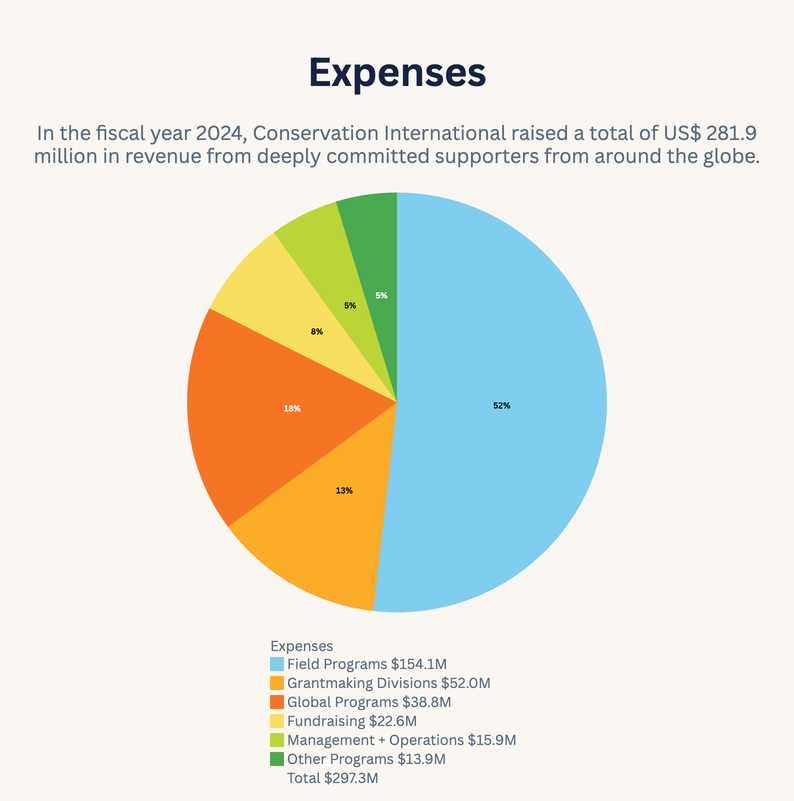
2024 Annual Report
These are unsettling and uncertain times, to say the very least. But that should only reinforce our directive: If we hold out for certainty, we will always be a step behind. Conservation was — and still is — a crisis discipline. That means we must be bold; try things that haven’t been done; trust science; and listen to, and learn from, those communities who have respected their relationship with the natural world.”
Peter Seligmann, Chairman of the Board
Highlights from 2024
(Download PDF at bottom of page for more stories from the field)
Nature for Climate
Small town, big impact
With support from Conservation International, the tiny town of Sena passed a law protecting 450,000 hectares (1.1 million acres) of remote lowland forest in northwestern Bolivia.
Over the past 25 years, Bolivian towns like Sena have protected a combined 100,000 contiguous square kilometers (38,600 square miles) of Bolivia’s Amazon — an area nearly the size of Iceland — securing livelihoods, protecting endangered wildlife and helping the country reach its goal to protect 30 percent of its land years ahead of schedule.
Scientists have said that 80 percent of the Amazon basin needs to be conserved. With about half already under some form of legal conservation status, much progress has been made, but much remains. Conservation International’s ambitious goal: protect 200,000 square kilometers (77,200 square miles) by supporting the creation of new conservation areas and the titling of Indigenous territories.
“Piece by piece, we are knitting together the fabric of conservation in the Amazon,” said Eduardo Forno, vice president of Conservation International-Bolivia. “Local communities have kept their eyes on the prize. They are having a big impact on the Amazon — for the benefit of us all.”
A new ‘wave’ of conservation?
Surf breaks aren’t just prized for their beauty and epic waves — they’re also a powerful force against climate change.
Thousands of surf breaks worldwide are surrounded by ecosystems that store massive amounts of potentially planet-warming carbon, according to new research by scientists from Conservation International and others.
Researchers mapped more than 4,800 popular surf spots across 113 countries and found that they store the same amount of carbon as the emissions from 77 million gas-powered cars.
“Our findings strengthen the case for protecting surf breaks and the surrounding ecosystems,” said Scott Atkinson, Conservation International’s surf conservation lead. This research can help motivate governments to create what are known as “Surf Protected Areas” — to date, Conservation International has helped create 23 such areas in Indonesia and is working to strengthen more in Costa Rica, Peru and Brazil.
Ocean Conservation atScale
In Indonesian waters, a new debt swap makes waves
Last year, the government of Indonesia announced a deal to redirect more than US$ 35 million it owes to the United States into the conservation of coral reefs in the most biodiverse ocean area on Earth.
The swap will fund coral restoration in two key areas of the Pacific Ocean’s Coral Triangle that encompass three-quarters of the world’s coral species and more than 3,000 species of fish, turtles, sharks, whales and dolphins.
Conservation International worked on enabling the deal and has committed to furnishing US$ 3 million to support the debt swap. While Indonesia has participated in three earlier swaps, this will be the world’s first to focus on protecting coral reefs.
In 1987, Conservation International pioneered the first-ever “debt-for-nature” swap, enabling the government of Bolivia to retire US$ 650,000 in foreign debt (US$ 1.8 million in today’s dollars) in exchange for establishing three protected areas near the headwaters of the Amazon. Groundbreaking at the time, such debt swaps now are a mainstay of conservation.
“We never imagined that this critical mechanism to alleviate burdensome debt and protect the world’s most valuable ecosystems would eventually unlock billions for global conservation," said Conservation International CEO M. Sanjayan after the deal was announced.
Expedition finds trove of potentially undiscovered species
Around our blue planet, a raft of new discoveries provided a powerful reminder that there is much that our species does not know about what lies beneath the waves.
A recent deep-sea expedition off the coasts of Chile and Peru is revealing the secrets of a vast underwater mountain system — and making the strongest case yet for greater protections there.
Using underwater robots capable of descending more than 4,500 meters (14,760 feet), researchers say they may have discovered more than 100 never-before-seen species living on the Salas y Gómez and Nazca ridges, which stretch across the southeastern Pacific. The monthlong expedition, led by the Schmidt Ocean Institute — with funding from Conservation International, the Blue Nature Alliance and Coral Reefs of the High Seas Coalition — explored 10 seamounts, some for the first time.
Ocean advocates hope a high seas treaty approved by the United Nations in 2023 will help create new marine protected areas to shield international waters from increasing threats — arguing that the Salas y Gómez and Nazca ridges should be one of the first areas considered for protection.
Regenerative Economies
An ancient tradition is saving an African grassland
Few places on Earth are as evocative as the grasslands of sub-Saharan Africa — nor host as much iconic wildlife. Thanks to Conservation International, these places are being brought back to health.
In South Africa, people long raised livestock alongside wildlife, mimicking the rhythms of nature. But when apartheid arrived, centuries of traditional herding were disrupted when communities were forcibly displaced from their ancestral territories, reshaping their cultural identity and way of life.
Working alongside pastoral communities, our local affiliate, Conservation South Africa, is restoring these vital grasslands, in part by reinvigorating a herding approach that had been practiced here for thousands of years. This effort aims to protect more than 30,000 hectares (74,000 acres) of land while establishing a replicable model to conserve grasslands throughout Africa.
Through the project, communal herders agree to move livestock periodically between different pastures, allowing grazed lands to recover. In exchange, farmers receive incentives such as vaccinations for their cattle and opportunities to sell their cattle to prime buyers. This “conservation agreement” model — developed by Conservation International 20 years ago — is reaping benefits for herders: As a result of the project, livestock have more to eat and arrive at market healthier, fatter and much more likely to command a premium price.
“This is a way of doing things that honors the whole system — people, livestock, wildlife and plants all thriving together,” said Julia Levin, who leads Conservation South Africa. “But it’s important to remember that no one invented this model — this is simply what African pastoralism looks like in its most innate form.”
How a predator helped bring one community back to life
When a small town saw its livelihood drying up, it had a choice — find new ways to make money, or overcome its fears and enlist an unlikely ally: Crocodiles.
About a decade ago, the small fishing community of Topón in southern Mexico noticed that the shrimp on which its economy depended were dying off. It took time to determine that one of the factors behind the decrease in shrimp populations was the elimination of the estuary’s native crocodiles, which were feared — and often hunted — by the community.
A project implemented by Conservation International and partners sought to change that.
Crocodiles are wetland engineers, said Conservation International’s Ramón Flores. Their swimming movements stir up sediment, aerating the water, improving oxygen levels and keeping minerals and nutrients from settling. “Without the crocodile, the wetland ceases to exist,” Flores said. “If you want good fishing, crocodiles must be present. Their decline is one of the main reasons that shrimp numbers plummeted.”
So the project hosted workshops that showed the community the critical role that crocodiles play in keeping the ecosystem healthy — and ensuring there is an abundance of shrimp to fish. The project also organized monitoring brigades to keep an eye on the species and measure progress — an eye-opening experience for the community, Flores said.
For the community, “witnessing the crocodile’s life cycle play out in front of them had a profound and moving effect,” he said. As for shrimp, the fishermen’s catch has increased tenfold since the project began more than five years ago. The local economy is also showing signs of improvement, notably for women and youth. With a new shrimp processing plant, the fishermen hope to connect directly to higher-value markets that support sustainable production.
“None of this would have been possible without taking this holistic and community-driven approach,” Flores said. “These are essential pieces to long-term sustainability.”
Science and FinanceInnovations
To restore grasslands, it’s time to get wild
Stretching across 40 percent of the planet, grasslands hold more than a third of the world’s land-based carbon in their vast underground root and soil systems.
They are bastions of biodiversity and contribute to the livelihoods of a billion people worldwide. Yet in some cases, grasslands are disappearing faster than forests.
“Ironically, some grasslands are being threatened by inappropriate climate actions, like planting trees where they don't belong,” said Conservation International scientist Heidi-Jayne Hawkins. “That’s a problem for all grassy ecosystems — so, we asked, what can we do that’s really a win for people, biodiversity and climate?”
According to a study co-led by Hawkins, one answer lies in bringing wildlife back to the grassy ecosystems they have helped shape for millennia — or at least mimicking the natural processes that keep grasslands in balance.
The study found that returning savannas to wilder states has clear benefits. Restoring populations of wild animals to historical levels keeps lands healthy. Healthier lands, in turn, are good for the climate and communities — including for ranchers’ and farmers’ livelihoods.
New study links birds’ diets to forest health
When you’re hungry and far from home, you might find yourself being less picky about what you eat.
Not so for birds. The farther they are from the core of their habitats, the pickier they get, according to a first-of-its-kind study from Conservation International and others that investigated the feeding behavior of nearly 100 species.
Why it matters: Birds disperse the seeds of about 90 percent of all tropical plants.
The study found that birds’ feeding preferences are highly connected to environmental stressors — and could have major impacts on ecosystems, says Conservation International climate expert Camila Donatti, a study co-author.
“In general, birds look for fruits that perfectly fit the size of their beak — so they can just grab and go,” she said. “We thought that in the birds’ core habitat, they would be choosier about the fruit they eat since there is a larger variety of it.”
But Donatti and others found that birds at the edges of their habitats contend with different climate conditions, different predators or competition for resources. “Being at the outer limits of their habitats is stressful, so they have to be careful to eat the exact plants that give them the biggest return on investment,” she said. “They get pickier because they don’t have energy to waste.”
This is significant because climate change and human activities are pushing bird species outside their range, creating a mismatch between the types of birds in a particular area and the food available to them.
“If these systems get out of balance, they won’t function in the way we expect them to and provide the essential services we rely on, from food to clean water to climate regulation,” Donatti said. “If we are going to protect that, we have to understand how it works.”
In Kenya, a global crisis sparks a new era of conservation
One of the most iconic landscapes in the world, Kenya’s Maasai Mara, thrives thanks to tourism income.
When COVID halted global travel in 2020, communities in the region that lease their land to wildlife conservancies and tour companies weighed selling some of it, risking the habitats that sustain the very wildlife that draw tourists.
But from this crisis was born a new way of doing conservation.
In 2020, Conservation International and the Maasai Mara Wildlife Conservancies Association launched the African Conservancies Fund — a rescue package to offset lost revenues for thousands in the area who rely on tourism income.
The conservancy model enables people living near national parks to combine their properties into large protected areas and earn income by leasing that land for safaris and lodges. Communities in Maasai Mara have now created 24 conservancies, protecting a total of 180,000 hectares (450,000 acres) — effectively doubling the total area of habitat for wildlife in the region.
But elsewhere in Africa, the conservancy model has remained out of reach.
“Conservancies have the potential to lift pastoral communities out of poverty in many African landscapes. But starting a conservancy requires significant funding — money they simply don’t have,” said Bjorn Stauch, who leads Conservation International’s conservation finance division.
Now, Conservation International is finding a way for communities to start conservancies and strengthen existing ones. Over the next three years, the organization aims to invest millions of dollars in new and emerging conservancies across southern and eastern Africa.
Loans issued by the fund — now called the African Conservancies Facility — will be repaid through tourism leases, jumpstarting new conservancies and reinforcing those already in place.
Keep Reading
Download Annual Report
Read what is next in 2025 — you will see our tireless effort to live up to our ideals — creating breakthrough ideas that protect and restore nature for people.